炎症性肠病(IBD)主要指三种不同的临床表现:克罗恩病、溃疡性结肠炎和眼袋炎。这些疾病的特点是胃肠道不同程度的持续性黏膜炎症。一般来说,IBD表现出波动性活动,伴有一阵阵不受控制的慢性黏膜炎症,随后在缓解期发生重塑过程。在过去50年中在西欧这类疾病的发病率稳步增长,现在在亚洲和东欧国家迅速扩大。IBD也正在成为年轻人的一个重要负担。
这些慢性炎症的确切病因仍有待阐明,但导致粘膜炎症病变的最重要的病理生理机制正在被揭示。
这些机制是环境、遗传和免疫调节因子复杂相互作用的结果。肠道菌群和黏膜免疫系统之间的异常通讯被认为是导致遗传易感个体IBD的核心缺陷。在胃肠道内,共生菌的炎症能力各不相同。一些寄居细菌是促炎性的,而其他细菌则减弱炎症反应。菊粉和低聚果糖等益生元可通过增加有益健康的细菌数量和活性来改善人体肠道菌群的平衡。本文综述了支持使用益生元预防和控制IBD的实验和临床证据。
肠道菌群
“微生物区系”或“微生物群落”是指在寄主个体的特定生态位中聚集的活微生物群落。人类肠道是大量、多样和动态的微生物种群的自然栖息地,这些微生物在几千年前已经适应了生活在黏膜表面或腔体内。小肠内的细菌数量增加,从空肠中的大约104个增加到回肠远端每克管腔中107个菌落形成单位。大肠是肠道中细菌最密集的区域,那里有几百克细菌,肠腔内含物密度为每克含1012个菌落形成单位。
我们目前对健康和疾病中肠道生态系统微生物组成的了解仍然非常有限。使用传统微生物培养技术的研究只能回收一小部分粪便细菌。通过粪便标本显微镜检查发现,超过50%的细菌细胞不能在培养物中生长。基于细菌基因组序列多样性的分子生物学技术正被用来描述不可培养的细菌。对粪便微生物群的分子研究表明,在人类肠道生态系统中,只有55个已知的“细菌”分支或超级界中有7个被检测到,其中3个细菌分支占主导地位,即类杆菌、硬壁菌和放线杆菌。然而,在物种和菌株水平上,个体之间的微生物多样性是非常显著的。每个人都有自己独特的细菌组成模式。
另一方面,将在无菌条件下繁殖的动物与常规饲养的动物进行比较,研究结果清楚地表明了寄居细菌对宿主生理的重要影响。肠道细菌与宿主之间的相互作用是一种对双方都有利的共生关系。寄主提供了一个营养丰富的栖息地,细菌给寄主带来了重要的好处。微生物群的功能包括营养(发酵不易消化的底物,产生短链脂肪酸、吸收离子、产生氨基酸和维生素)、保护(防止外来微生物入侵的屏障效应),以及对肠上皮和免疫系统的营养作用(局部和系统免疫的发展和稳态)。
在无菌环境中培育的动物,肠道黏膜中淋巴细胞密度低,血清免疫球蛋白浓度低。接触共生微生物后,粘膜淋巴细胞数量迅速增加,淋巴滤泡中生发中心的大小也随之增大。免疫球蛋白在血清中含量明显增加。最有趣的是,最近的研究结果表明,一些共栖体在诱导肠道淋巴滤泡中调节性T细胞中起主要作用。调节性T细胞介导的调节途径是重要的稳态机制,通过这种机制,宿主可以耐受肠道内或其他身体表面的大量无害抗原,而不会通过炎症反应。
细菌与炎症性肠病
IBD患者肠道菌群中细菌抗原的调节和耐受机制似乎发生了改变。正常的黏膜防御主要基于分泌到肠腔中的IgA抗体,并中和肠腔中的微生物,从而避免粘膜炎症。然而,在IBD中,黏膜产生的抗肠道细菌IgG抗体显著增加,黏膜防御依赖于组织内IgG介导的反应和组织中高活化的淋巴细胞固有层对细菌抗原反应。这些事件会导致炎症和组织损伤。这种改变的免疫反应并不是针对一组潜在的病原体,而是涉及大量和未确定数量的属于常见肠道菌群的共生物种。肠道生态系统中的微生物失衡可以解释黏膜免疫系统对肠道细菌的异常反应。
一些研究表明,IBD患者和健康对照者的粪便微生物群组成不同。分子研究表明,在活动性克罗恩病或溃疡性结肠炎患者中,相当大比例的粪便细菌(占优势菌群的30%至40%)属于在健康受试者中不常见的系统发育群。这些显著的变化可能是继发于疾病活动,但在感染性腹泻患者中没有观察到。另一方面,研究表明IBD患者粪便和粘膜相关群落中细菌种类的多样性降低。Manichanh及其同事采用了一种宏基因组方法对克罗恩病的细菌多样性进行了详尽的调查,发现缓解期患者的硬壁菌数量与健康对照组相比显著减少(图1)。
对粘膜相关细菌的研究发现,在临床活动期溃疡性结肠炎或克罗恩病患者中有高浓度的粘附细菌,但在健康对照组中没有。随着黏膜炎症程度的加重,粘膜粘附细菌的浓度逐渐增加,所鉴定的细菌来源于粪便。荧光原位杂交(FISH)技术显示溃疡性结肠炎和克罗恩病患者的大多数粘膜标本中有细菌侵入粘膜,但对照组的任何粘膜标本中均未发现细菌侵袭。入侵细菌属于多种属,包括蛋白细菌、肠杆菌科、类杆菌/普雷沃特菌群、梭状芽孢杆菌和硫酸盐还原菌。然而,没有检测到双歧杆菌或乳酸杆菌侵入粘膜。此外,Macfarlane及其同事观察到溃疡性结肠炎患者直肠活检中粘附的非侵袭性双歧杆菌数量低于对照组。
益生元
一个健康或“平衡”的微生物群被认为是一个以糖分分解为主,包含大量双歧杆菌和乳酸杆菌的微生物群。菊粉和低聚果糖是益生元碳水化合物,在人体胃肠道中抵抗肠道和胰腺酶的消化,并由生活在肠道生态系统中的细菌发酵。当给予足够的量时,这些益生元会增加肠道内的糖化活性,并选择性地促进双歧杆菌的生长。大量研究表明,食用菊粉或低聚果糖的受试者粪便中双歧杆菌数量增加。此外,口服菊粉和低聚果糖的摄入量增加。
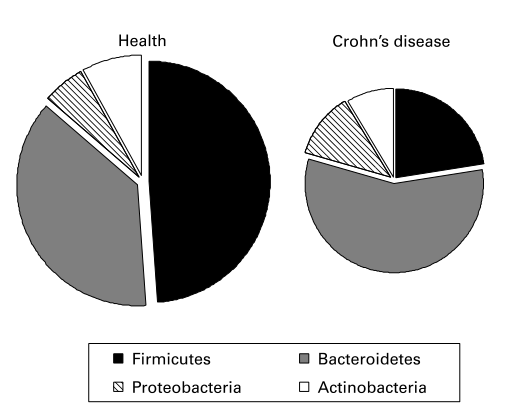
图1. 克罗恩病患者的粪便微生物群中含有的硬壁菌比例降低。该图显示了来自Manichanh及其同事(参考文献18)的数据,并代表了6名健康人和6名临床病情缓解患者的每个部门的系统类型数。
人类结肠粘液相关群落中双歧杆菌和乳酸杆菌的数量。Langlands等人表明,在喂食含菊粉和低聚果糖的益生元混合物2周的受试者中,近结肠和远端结肠的活检粘膜标本中双歧杆菌和乳酸杆菌的数量可以增加10倍以上。同样,一项针对溃疡性结肠炎患者的研究显示,使用双歧杆菌菌株和富含低聚果糖的菊粉制成的合生元制剂后,直肠黏膜上的双歧杆菌数量增加了。
假设通过增加黏膜表面的“友好”细菌数量,菊粉和低聚果糖可以改善IBD中的屏障功能,并阻止能够侵入的需氧肠道细菌的粘膜定植。这一假设已经在大量的实验研究中得到验证,这些实验研究使用了不同的IBD动物模型。
炎症性肠病的实验模型
在化学右旋糖酐硫酸钠(dss28)诱导的大鼠结肠炎模型中检测了益生元菊粉的作用。口服DSS超过3至5天可诱导对结肠上皮细胞的直接毒性,导致粘膜屏障功能障碍,增加对大分子的通透性。这些事件之后是隐窝破坏和肠绒毛高度降低,随后是细菌入侵和黏膜炎症。在大鼠中,每天口服菊糖可增加盲肠腔中的本地乳酸杆菌数量,并降低结肠内pH值。在暴露于DSS诱发结肠炎的大鼠中,口服菊糖可显著降低组织髓过氧化物酶活性,这是中性粒细胞浸润的一个指标,以及炎症介质的黏膜释放。此外,菊粉喂养的大鼠表现出粘膜损伤程度的减轻和隐窝破坏的严重程度的减轻。菊糖治疗组大鼠的组织学损伤评分明显低于对照组(图2)。口服菊粉治疗无论是在接触DSS之前还是在接触DSS期间都是同样有效的。
低聚果糖和菊粉单独或与益生菌双歧杆菌联合使用的效果最近在同一DSS模型中进行了测试。益生元单独或与婴儿双歧杆菌联合使用可显著改善疾病活动性指标,降低结肠髓过氧化物酶活性和炎症介质的表达。有趣的是,与结肠炎对照组相比,益生菌、益生元或两者联合治疗的大鼠肠系膜淋巴结和肝脏的细菌移位明显减少。作者的结论是,低聚果糖和菊粉以及双歧杆菌菌株可以防止细菌入侵,并在该模型中具有抗炎作用。
在远端结肠中使用橡胶套管,单次结肠内注射稀释在乙醇中的三硝基苯磺酸(TNBS)(通常20-50mg TNBS:30-50%乙醇)可诱发远端结肠的慢性炎症性病变。低聚果糖的作用已在结肠炎的TNBS模型中进行了测试。口服低聚果糖显著降低TNBS治疗大鼠的结肠内pH值、宏观损伤评分和组织髓过氧化物酶活性。此外,低聚果糖增加了盲肠内容物中乳酸和丁酸的浓度以及乳酸菌的数量。在随后的辅助实验中,这些研究者证明了直接将乳酸菌与短链脂肪酸一起灌注是重现低聚果糖抗炎作用的必要条件。他们得出结论,乳酸菌发酵益生元是介导抗炎作用的主要机制。
进一步的实验工作评估了菊糖和低聚果糖在转基因HLA-B27大鼠自发性结肠炎模型中的抗炎作用。转基因人类HLA-B27-beta2微球蛋白基因的大鼠在2-4个月大时会自发发生严重程度不同的免疫介导性结肠炎。该病的特点是非血性腹泻,盲肠和结肠黏膜有明显的炎性浸润。Hoentjen和他的同事在这个自发性结肠炎模型中测试了低聚果糖和菊粉的混合物,并观察到益生元混合物喂养的大鼠具有显著的抗炎作用。益生元治疗可降低病变的大体形态学评分和组织学分级。此外,益生元治疗降低了促炎细胞因子如IL-1b的表达,但促进调节型细胞因子(TGF-b)的表达。
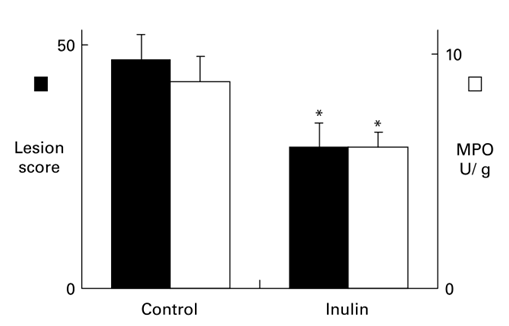
图2. DDS诱导的大鼠结肠炎结肠黏膜病变评分(实心柱,左y轴)和组织中髓过氧化物酶(MPO,开放柱,右y轴)含量。每日经口灌胃给予400毫克菊糖可显著降低结肠组织的病变评分和髓过氧化物酶含量(见参考文献28)。
益生元乳果糖的作用也在一些肠道炎症动物模型中进行了测试。缺乏IL-10基因的小鼠会自发地发展成结肠炎。在新生期,这些基因敲除小鼠结肠中的乳酸菌种类减少,粘附和转位细菌增加。口服乳果糖可使粪便中的乳酸杆菌计数正常化,并防止结肠炎的发生。同样,乳果糖的保护作用已在DSS和TNBS模型中得到证明。综上所述,所有这些实验数据有力地表明了益生元在各种IBD动物模型中的抗炎作用。
临床研究
一项随机、安慰剂对照、双盲、交叉临床试验测试了菊粉对慢性眼袋炎患者的疗效。这种临床状况的特点是慢性黏膜炎症的回肠袋肛管吻合术的病人,已进行了全结肠切除。回肠袋是通过外科手术构建的,以便起到粪便贮存器的作用。炎症性疾病损害了储液罐的功能,导致持续腹泻,伴有粘液和血液。20名有轻度疾病活动的患者进入试验,随机开始服用安慰剂或菊粉(每天24克),为期3周,采用双盲交叉设计,洗出期为4周。与安慰剂相比,饮食中添加菊粉可显著降低回肠贮存器黏膜炎症的内镜和组织学参数(表1)。这种影响与粪便中丁酸盐的增加和粪便中类杆菌数量的减少有关。
Furrie等27报道了一项随机、安慰剂对照、双盲临床试验,对两组平行的溃疡性结肠炎患者进行了研究。符合条件的患者有轻微的疾病活动,并且正在服用稳定的药物。18名患者被随机分为1个月,接受合生元制剂(每天12克低聚果糖强化菊粉和每天2000亿个菌落形成单位的长双歧杆菌)或安慰剂(麦芽糊精)。合生元治疗可显著降低促炎性细胞因子(TNF-α、IL-1b)和诱导β防御素的黏膜表达。活检组织学检查显示,接受合生元治疗的患者炎症细胞浸润和隐窝脓肿明显减少,乙状结肠镜评分和临床活动指数也有所改善,但由于登记的患者人数减少,差异不显著。
富含低聚果糖的菊粉对活动性溃疡性结肠炎患者的疗效最近在一项随机、安慰剂对照、双盲试验中进行了试验,试验对象为两个平行组。符合条件的患者以前曾使用美沙拉秦作为维持治疗或不使用药物缓解,并因轻度-中度活动复发而到医院就诊。他们接受美沙拉秦(3g/天)治疗,并随机分配接受富含低聚果糖的菊粉(12g/天)或安慰剂(12g/天麦芽糊精)两周。主要终点是益生元的抗炎作用,通过客观的、非侵入性的肠道炎症标志物(即钙卫蛋白的粪便浓度)进行评估。钙保护蛋白是一种存在于粒细胞中的蛋白质,可抵抗代谢降解,可在粪便中测定。有趣的是,在第7天,在服用富含低聚果糖的菊粉组中观察到钙保护蛋白的早期显著降低,而安慰剂组则没有。在研究期结束时,两组患者的疾病活动评分显著降低。使用这种益生元可以提高对美沙拉秦药物治疗的反应,但这一点需要在有足够数量患者的试验中进一步研究。
益生元在克罗恩病中也得到了检测。在一项小型开放性试验中,10名活动性回肠-结肠克罗恩病患者连续3周每天服用15克低聚果糖。口服低聚果糖三周后,除两名患者外,所有患者的Harvey-Bradshaw疾病活动指数均下降,与基线相比,整个组的疾病活动性显著下降。粪便中双歧杆菌数量显著增加,但直肠活检中没有。然而,这项研究不包括安慰剂对照组。目前,同一组研究人员正在对克罗恩病患者进行一项具有适当样本量的对照研究。
综上所述,迄今为止的实验和临床数据支持这样一个假设,即菊粉和低聚果糖等益生元可以为预防或减轻人类克罗恩病、溃疡性结肠炎和眼袋炎的肠道炎症病变提供机会。对照临床试验我们还需要适当的样本量来证实这个假设。
表1 日粮中添加菊粉对眼袋炎活动指数(PDAI)的影响
| 安慰剂 | 菊粉 | P |
临床评分大便次数 | 0.53(0.19) | 0.47(0.15) | 0.65(NS) |
直肠出血 | 0.05(0.06) | 0.05(0.06) | 0.10(NS) |
大便急迫/腹部绞痛 | 0.68(0.18) | 0.47(0.19) | 0.16(NS) |
发烧 | 0(0) | 0(0) | 0·10(NS) |
临床总酸痛 | 1.26(0.29) | 1.00(0.27) | 0.17(NS) |
内镜评分水肿 | 0.16(0.09) | 0(0) | 0.08(NS) |
粒度 | 0.05(0.06) | 0.05(0.06) | 1.00(NS) |
易碎性 | 0.32(0.12) | 0.16(0.09) | 0.18(NS) |
血管丧失 | 0.05(0.06) | 0.11(0.08) | 0.56(NS) |
粘液渗出物 | 0.32(0.12) | 0.05(0.06) | 0.03 |
溃疡 | 0.58(0.12) | 0.58(0.12) | 1.00(NS) |
内镜总评分 | 1.47(0.32) | 0.95(0.22) | 0.04 |
组织学评分 多晶渗透 | 1.44(0.15) | 1.11(0.14) | 0.05(NS) |
每低倍视野溃疡 | 1.17(0.13) | 1(0) | 0.18(NS) |
组织学总评分 | 2.61(0.26) | 2.11(0.14) | 0.04 |
PDAI总分 | 5.39(0.62) | 4.05(0.44) | 0.01 |
数据是平均值和标准误差的平均值,在括号内,由韦尔斯和同事发表(参考文献36)。NS=不显著。


